Finding Conserved Quantities in a Pathway Model
This example shows how to use the sbioconsmoiety function to find conserved quantities in a SimBiology® model.
Aims
Use
sbioconsmoietyto determine the conserved quantities present in two models of glycolysis in T. brucei.Use the computed conserved quantities in an analysis of these models.
Background
Trypanosoma brucei is the single-celled, eukaryotic parasite responsible for African sleeping sickness. This organism survives inside an infected host by metabolizing glucose from the host bloodstream. In T. brucei as well as other trypanosomes, a large portion of glycolysis occurs inside a specialized organelle called the glycosome.
To investigate the function of the glycosome, Bakker et al. (2000, 1997) constructed and validated a computational model of glycolysis in T. brucei that explicitly includes glycosomal compartmentation. They compared the properties of this model to those of a derived model in which the glycosome is absent. Among other results, they found that in the absence of the glycosome, hexose phosphate intermediates in the glycolytic pathway can accumulate to high levels that would be hazardous to the cell. In their analysis, Bakker et al. were able to explain how the compartmentation of metabolites provided by the glycosome prevents this potentially toxic accumulation.
One way to understand the effect of compartmentation is to examine how it affects the conserved quantities present in the system. In this example, we calculate the conserved quantities in the two models of T. brucei glycolysis and discuss their significance in the context of the analysis of Bakker et al.
Load the Project
Begin by loading the project at the command-line using sbioloadproject.
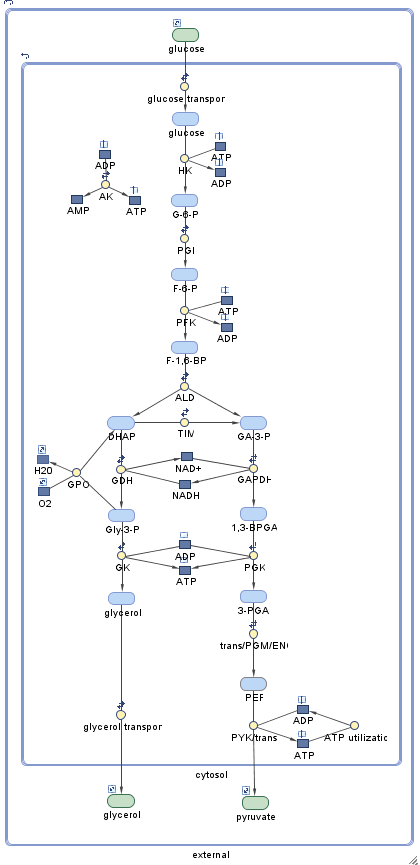
sbioloadproject trypanosome_glycolysisThe project contains two models. The first model, m1, contains the wild-type glycolysis network displayed below. (You can explore the network interactively by starting the SimBiology Model Builder app with simBiologyModelBuilder and opening the project file trypanosome_glycolysis.sbproj located in (matlabroot/toolbox/simbio/simbiodemos.)

This system is a slightly simplified version of the pathway used by Bakker et al. The model has three compartments: glycosome, cytosol, and external. Metabolites contained in the glycosome are in blue, while metabolites in the cytosol or external to the cell are in green. Some species, such as glycerol 3-phosphate (Gly-3-P), are present in multiple compartments.
The pathway begins with the import of extracellular glucose into the glycosome (for convenience, the cytosol is "skipped" in this process). The pathway proceeds downwards in the diagram, ending with the transport of pyruvate out of the cytosol. Under aerobic conditions, glycerol 3-phosphate (Gly-3-P) is oxidized via glycerol-3-phosphate oxidase (GPO) outside the glycosome; as a consequence, glucose is fully converted to pyruvate. Under anaerobic conditions, this reaction does not occur and the glycolytic pathway produces glycerol in addition to pyruvate.
Compute the Conserved Quantities in the Wild-Type Network
The function sbioconsmoiety examines the structure of a model's stoichiometry matrix to find linear combinations of species that are conserved. This analysis is structural in that it relies only on the stoichiometry and structure of the network and not on reaction kinetics. In fact, all the reaction rates in this model have been set to 0 because these rates are not important to our analysis. Here we call sbioconsmoiety with the algorithm specification 'semipos', so that all conserved quantities returned involve only positive sums of species. The third argument 'p' asks for the output to be printed to a cell array of strings.
cons_wt = sbioconsmoiety(m1,'semipos','p')
cons_wt = 10×1 cell
{'external.glucose' }
{'external.glycerol' }
{'external.pyruvate' }
{'cytosol.H20' }
{'cytosol.O2' }
{'cytosol.Gly-3-P + cytosol.DHAP' }
{'cytosol.ATP + cytosol.ADP + cytosol.AMP' }
{'glycosome.ATP + glycosome.ADP + glycosome.AMP' }
{'glycosome.NAD+ + glycosome.NADH' }
{'2 glycosome.ATP + glycosome.ADP + glycosome.G-6-P + glycosome.F-6-P + 2 glycosome.F-1,6-BP + glycosome.DHAP + glycosome.GA-3-P + glycosome.Gly-3-P + glycosome.1,3-BPGA'}
The last cell in the cell array contains a long string. Break this string up and display it so it can be read.
disp(cons_wt{end}(1:68));2 glycosome.ATP + glycosome.ADP + glycosome.G-6-P + glycosome.F-6-P
disp(cons_wt{end}(69:147));+ 2 glycosome.F-1,6-BP + glycosome.DHAP + glycosome.GA-3-P + glycosome.Gly-3-P
disp(cons_wt{end}(148:end));+ glycosome.1,3-BPGA
The output of sbioconsmoiety contains ten quantities whose time rate of change is zero, regardless of reaction kinetics. There are two conserved pools of the adenine nucleotides ATP, ADP, and AMP, one in the glycosome and one in the cytosol. The glycosomal pool of nicotinamide nucleotides NAD+ and NADH is conserved as well. The singly conserved species such as external.glucose and cytosol.O2 are species on the boundary of the system that have their BoundaryCondition property set to true. These species are included in the output of sbioconsmoiety because their amounts would indeed remain constant during a hypothetical simulation.
The remaining two conserved quantities represent pools of bound phosphate, one inside and one outside the glycosome. The one inside includes nine different species. Note that the coefficients of ATP and fructose-1,6-biphosphate (F-1,6-BP) are both 2, as these species each have two transferable phosphate groups.
The species participating in the conserved sum have been highlighted below. This figure was generated by selecting the relevant species in the Diagram Table View in the SimBiology desktop. The conserved cycle "begins" when glucose is phosphorylated by ATP to form glucose 6-phosphate (G-6-P). This phosphate group propagates down through the pathway until it is transferred back to ATP from 1,3-biphosphoglycerate (1,3-BPGA) or glycosomal glycerol 3-phosphate (Gly-3-P), completing the cycle.

Note that the sum cytosol.DHAP + cytosol.Gly-3-P arises as an independently conserved pool because the DHAP/Gly-3-P antiporter exchanges one glycosomal DHAP molecule for one cytosolic Gly-3-P molecule and vice versa. There are fluxes of phosphate groups in and out of this pool, but the next flux is zero because these fluxes cancel each other out.
View the Experimental Model with No Glycosome
Now let's consider the second model, m2, that contains the in silico experimental network of Bakker et al. in which the glycosome has been removed. In this model all metabolites reside in the cytosol. In particular, there is no longer an antiport exchange of DHAP and Gly-3-P in and out of the glycosome, and there is a single pool for the adenine nucleotides ATP, ADP, and AMP.
Compute the Conserved Quantities in the Experimental Network
cons_exp = sbioconsmoiety(m2,'semipos','p')
cons_exp = 7×1 cell
{'external.glucose' }
{'external.glycerol' }
{'external.pyruvate' }
{'cytosol.H20' }
{'cytosol.O2' }
{'cytosol.NADH + cytosol.NAD+' }
{'cytosol.AMP + cytosol.ADP + cytosol.ATP'}
The species on the boundary of the system are still present in the experimental model, and their amounts are again conserved. Without the glycosome, however, the conservation of bound phosphates has disappeared, leaving only conservation relations for the nicotinamide and adenine nucleotides.
Discussion
In their analysis of the function of the glycosome in T. brucei, Bakker et al. find that glycosomal compartmentation prevents the potentially toxic accumulation of the hexose phosphate intermediates G-6-P and F-1,6-BP during glycolysis. This observation can be understood in light of the observed difference in the conservation of phosphates with and without the glycosome. When the glycosome is present, intermediates such as G-6-P or F-1,6-BP cannot accumulate to arbitrarily high levels, as they are limited by the total amount of organic phosphate present in a conserved pool. Without the glycosome, this restriction is absent. Insight may also be gained by considering the glycosomal compartmentation of adenine nucleotides. When the extracellular level of glucose is increased, the reactions HK and PFK are stimulated. When the glycosome is present, these reactions are self-limiting, as they deplete the ATP from a conserved pool of glycosomal ATP, ADP, and AMP. When the glycosome is absent, on the other hand, the cytosolic ATP/ADP ratio in fact increases with increasing levels of extracellular glucose. As a consequence, the reactions HK and PFK are further stimulated, leading to the accumulation of their products, G-6-P and F-1,6-BP.
This analysis shows that glycosomal compartmentation provides a negative feedback mechanism on the buildup of intermediates. Bakker et al. suggest that the conserved pool of organic phosphates may also serve as an energy storage mechanism for wild-type T. brucei during times of starvation.
In this example we have shown how to calculate the conserved quantities in a SimBiology model and how an analysis of these conserved quantities can lead to insight into the behavior of a network.
References
Bakker, B. M., Mensonides, F. I. C., Teusink, B., van Hoek, P., Michels, P. A. M., and Westerhoff, H. V. Compartmentation Protects Trypanosomes from the Dangerous Design of Glycolysis. PNAS (2000) vol. 97, 2087-2092.
Bakker, B. M., Michels, P. A. M., Opperdoes, F. R., and Westerhoff, H. V. Glycolysis in Bloodstream Form Trypanosoma brucei Can Be Understood in Terms of the Kinetics of the Glycolytic Enzymes. J. Biol. Chem. (1997) vol. 272, 3207-3215.